
Introduction
The roots of about 90% of all plant families have species that are involved in mutualistic symbiotic rela-tionships with certain kinds of fungi. It has been estimated that these fungi probably amount to 15% of the total weight of the world’s plant roots. Associations of this kind are termed mycorrhizae, from the Greek words for fungus and root.
There are two principal types of mycorrhizae . In arbuscular mycorrhizae, the fungal hyphae penetrate the outer cells of the plant root, forming coils, swellings, and minute branches; they also extend out into the surrounding soil.
In ectomycorrhizae, the hyphae surround but do not penetrate the cell walls of the roots. In both kinds of mycorrhizae, the mycelium extends far out into the soil. A single root may asso-ciate with many fungal species, dividing the root at a millimeter-by-millimeter level.
Definition
The name mycorrhizas, which literally means fungus-root, was invented by Frank (1885) for non-pathogenic symbiotic associations between roots and fungi.
Mycorrhizas are symbiotic associations essential for one or both partners, between a fungus (specialized for life in soils and plants) and a root (or other substrate-contacting organ) of a living plant, that is primarily responsible for nutrient transfer.
Categories of Mycorrhizal Associations:
| No. | Category | Definition | Hosts | Fungi |
| 1 | Arbuscular mycorrhizas | Associations formed by Glomeromycotan fungi in plants that usually have arbuscules and often have vesicles (also known as vesicular-arbuscular mycorrhizas, AM, VAM). | Plants | Glomeromycota |
| 1.1 | Linear VAM | Associations that spread predominantly by longitudinal intercellular hyphae in roots (formerly known as Arum series VAM). | Plants | As above |
| 1.2 | Coiling VAM | Associations that spread predominantly by intracellular hyphal coils within roots (formerly known as Paris series VAM). | Plants | As above |
| 1.2.1 | Beaded VAM | Coiling VAM in roots, where interrupted root growth results in short segments divided by constrictions. | Woody plants | As above |
| 1.2.2 | Inner cortex VAM | Coiling VAM with arbuscules in one layer of cells of the root inner cortex. | Plants | As above |
| 1.2.3 | Exploitative VAM | Coiling VAM of myco-heterotrophic plants, usually without arbuscules. | Achlorophyllous plants | As above |
| 2 | Ecto-mycorrhiza (ECM) | Associations with a hyphal mantle enclosing short lateral roots and a Hartig net of labyrinthine hyphae that penetrate between root cells. | hosts | Higher fungi (asco-, basidio- and zygo- mycetes) – see Section 9 |
| 2.1 | Cortical | Hartig net hyphae penetrate between multiple cortex cell layers of short roots | Most are gymnosperm trees | As above |
| 2.2 | Epidermal | Hartig net fungal hyphae are confined to epidermal cells of short roots | Angiosperms (most are trees) | As above |
| 2.2.1 | Transfer cell | Epidermal Hartig net with transfer cells (plant cells with wall ingrowths) | Pisonia (Nyctaginaceae) | Tomentella spp. in Pisonia (Chambers et al. 2005) |
| 2.2.2 | Monotropoid | Exploitative epidermal ECM of myco-heterotrophic plants in the Ericaceae where individual hyphae penetrate epidermal cells. | Ericaceae (Monotropa, Pterospora, Sarcodes) | Basidiomycetes |
| 2.2.3 | Arbutoid | ECM of autotrophic plants in in the Ericaceae where multiple hyphae penetrate epidermal Hartig net cells. | Ericaceae (part only) | Basidiomycetes |
| 3 | Orchid | Associations where coils of hyphae (pelotons) penetrate within cells in the plant family Orchidaceae. | hosts | Most are basidiomycetes in Rhizoctonia alliance (listed below). |
| 3.1 | Orchid Root | Associations within a root cortex. | Orchidaceae | As above |
| 3.2 | Orchid Stem | Associations within a stem or rhizome. | Orchidaceae | As above |
| 3.3 | Exploitative Orchids | Associations of myco-heterotrophic orchids. | Orchidaceae (fully or partially achlorophyllous) | Orchid, ectomycorrhizal, or saprophytic fungi |
| 4 | Ericoid | Coils of hyphae within very thin roots (hair roots) of the Ericaceae. | Ericaceae (most genera) | Most are Ascomycetes (listed below) |
| 5 | Sub- epidermal | Hyphae in cavities under epidermal cells, only known from an Australian monocot genus. | Thysanotus spp. (Laxmaniaceae) | Unknown |
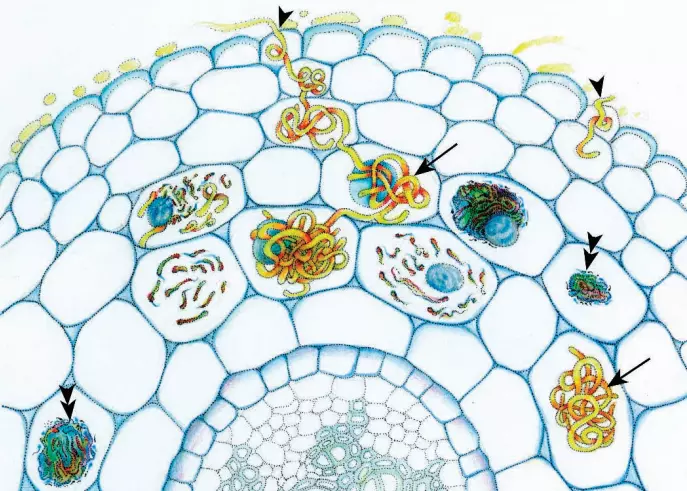
Arbuscular mycorrhizae
Arbuscular mycorrhizae are by far the more common of the two types, involving roughly 70% of all plant species (figure ). The fungal component in them are glomeromycetes, a monophyletic group that arose within one of the zygomycete lineages. The glomeromycetes are associated with more than 200,000 spe-cies of plants.
Unlike mushrooms, none of the glomeromycetes produce aboveground fruiting structures, and as a result, it is difficult to arrive at an accurate count of the number of extant species. Arbuscular mycorrh-izal fungi are being studied intensively be-cause they are potentially capable of increasing crop yields with lower phosphate and energy inputs.
The earliest fossil plants often show arbuscular mycorrhizal roots. Such associations may have played an important role in allowing plants to colonize land. The soils available at such times would have been sterile and lacking in organic matter. Plants that form mycorrhizal associations are particularly succes-sful in infer-tile soils; considering the fossil evidence, it seems reasonable that mycorrhizal associations helped the earliest plants succeed on such soils.
In addition, the closest living relatives of early vascular plants surviving today continue to depend strongly on mycorrhizae.
Some nonphotosynthetic plants also have mycorrhizal associations, but the symbiosis is one-way beca-use the plant has no photosynthetic resources to offer. Instead of a two-partner symbiosis, a tripartite symbiosis is established. The fungal mycelium extends between a photosynthetic plant and a non-
photosynthetic, parasitic plant. This third, nonphotosynthetic member of the symbiosis is called an epiparasite.
Not only does it obtain phosphate from the fungus, but it uses the fungus to channel carbohydrates from the photosynthetic plant to itself. Epiparasitism also occurs in ectomycorrhizal symbiosis.
Ectomycorrhizae
Ectomycorrhizae (see figure ) involve far fewer kinds of plants than do arbuscular mycorrhizae—perhaps a few thou-sand. Most ectomycorrhizal hosts are forest trees, such as pines, oaks, birches, willows, eucalyptus, and many others. The fungal components in most ectomycorrhizae are basidiomycetes, but
some are ascomycetes.
Most ectomycorrhizal fungi are not restricted to a single species of plant, and most ectomycorrhizal plants form associa-tions with many ectomycorrhizal fungi. Different combinati-ons have different effects on the physiological characteristics of the plant and its ability to survive under different environ-mental conditions. At least 5000 species of fungi are involved in ectomycorrhizal relationships.
Terminology
Symbiosis
It is the intimate associations between two or more different living organisms. Only a broad definition of symbiosis – living together of two or more organisms includes all types mycorrhizal associations (Lewis, 1985; Smith & Read, 1997).
Mutualism
A category of symbiotic associations where both partners benefit Mycorrhizas of myco-heterotrophic plants are not mutualistic as the fungus is being.
Mycorrhiza, Mycorrhizas, Mycorrhizal
These were defined by Frank (1885) as symbiotic associations between fungi and roots that are not pathogenic (i.e. intimate root-fungus associations without disease symptoms). Frank named these associations mycorrhizas which means fungus-root (they were formerly called mycorrhizae).
Myco-heterotrophic mycorrhizas
Non-mutualistic mycorrhizal associations where plants are parasitic on fungi (see Leake 1994). These plants are sometimes referred to as saprotrophic, cheating, myco-parasitic, etc. However, these should be referred to as myco-heterotrophic (fungus feeding) or exploitative associations.
Host Plant
A plant containing a fungus of any type.
Mycorrhizal fungi
These can be called a symbiont, associate, mycobiont, inhabitant, etc., but it is usually sufficient to call them fungi. Mycorrhizal fungi should not be called endophytes to avoid confusion with another major category of plant inhabiting fungi.
Colonisation
Neutral term ‘colonisation’ is preferential to infection (implying disease) when describing mycorrhizal fungus activity and the resulting fungal structures can be defined as colonies.
Inoculum
propagules of fungi capable of dispersing or initiating contact with plants.
Vascular plants
“Higher plants” with conducting elements for water and nutrients, differentiated leaves and roots, with a dominant sporophyte.
Roots
Plant organ responsible for nutrient uptake, mechanical support, storage, etc. that are usually subterranean.
Fungi
Members of the fungus kingdom are eukaryotic, heterotrophic organisms with a tubular body that reproduce by spores.
Mineral nutrients
The basic form of substances required for life (N, P, K, etc. excluding gases).
Photosynthesis
Solar energy capture by plants and conversion into organic carbon.
Root Terminology
Apoplast
The cell wall space inside living plants is collectively known as the apoplast.
Middle lamella
A cell wall zone rich in the carbohydrate pectin connecting adjacent cells.
Suberin
This is a hydrophobic material, containing lipids and phenolics, which impregnates the cell walls of specialised cells (Kolattukudy 1984). Suberin is thought to prevent the passage of water and other materials in the apoplast.
Suberin lamellae
These are concentric layers of suberin deposited on the inner surface of cell walls and considered to function as barriers to microbial and solute penetration. These are most often found in endodermal or exodermal cells.
Mucilage
High molecular weigh, poorly diffusible substances actively secreted by root epidermal and root cap cells (Rougier & Chaboud 1985). These primarily consist of carbohydrates, but also may contain sloughed cells, enzymes, phenolic compounds, etc.
Exudates
Root exudates are defined as substance released into the substrate by healthy and intact plant roots (Rovira 1969). These include water, sugars, amino acids, etc.
Exodermis
The hypodermis is called an exodermis if its cell walls contain a Casparian band and these cells often also have suberin lamellae (Peterson 1988).
Aerenchyma
Air spaces within plant organs. These can form between cells, or in the case of large spaces result from cell death. They often form continuous channels along the length or organs such as the root. The main role of aerenchyma is to provide gas exchange to cells in waterlogged soils (Armstrong 1979).
Periderm
The bark layer formed on the surface of roots or branches by secondary growth. Walls of periderm cells are strengthened by suberin and lignin deposits, which reduce their permeability and susceptibility to microbial activity and adverse soil conditions.
Metacutinization
This is the modification of dormant root tips by suberisation of one or more root cap cell layers (Romberger 1963). Inactive roots of many perennial plants develop a metacutinized apex, which functions as an extension of the exodermis, presumably for protection from adverse soil factors (Brundrett etc).
Rhizosphere
The zone surrounding roots where soil properties and microbial populations are influenced by root exudates.
Rhizoplane
The surface of the root and habitat for organisms which live in contact with the root.
References:
Mycorrhizae Book By Smith 1997
Mycorrhizael Anatomy and molecular biology Book by Patel
Raven biology book