
In This Topic, we discuss a detailed account of the ORIGIN OF ANGIOSPERMS. Angiosperms from the most dominant group of plants with at least 253,300 species (Thorne, 2007). Angiosperms are also found in a far greater range of habitats than any other group of land plants.
The phylogeny of angiosperms is a much-debated subject, largely because of very poor records of the earliest angiosperms. These earliest angiosperms probably lived in habitats that were not best suited for fossilization.
- Origin of Angiosperms
- What are Angiosperms?
- What is the age of Angiosperms?
- What is the Place of their Origin?
- Are Angiosperms Monophyletic or Polyphyletic?
- What are the possible ancestors of angiosperms?
- Euanthial Theory
- Pseudanthium Theory
- Anthocorm Theory
- Transitional-Combinational Theory
- Origin of Monocotyledons
- Questions about the Origin of Angiosperms:
- Sources and References:
Origin of Angiosperms
With the exception of conifer forest and moss-lichen tundra, angiosperms dominate all major terrestrial vegetation zones, account for the majority of primary production on land, and exhibit extensive morphological diversity.
Unfortunately, much less is known about the origin and early evolution of angiosperms, resulting in a number of different views regarding their ancestors, the earliest forms, and the course of evolution. The origin of angiosperms may be conveniently discussed under the following considerations.

What are Angiosperms?
Angiosperms form a distinct group of seed plants sharing a unique combination of characters. These important characteristics include carpels enclosing the ovules, pollen grains germinating on the stigma, sieve tubes with companion cells, double fertilization resulting in the triploid endosperm, and highly reduced male and female gametophytes.
The angiosperms also have vessels. The pollen grains of angiosperms are also unique in having non-laminate endexine and ectexine differentiated into a foot layer, columellar layer, and tectum.
The angiosperm flower typically is a hermaphrodite structure with carpels surrounded by stamens and the latter by petals and sepals, since insect pollination prevails.
Arbuscular mycorrhizae are also unique to angiosperms. The vessel elements of angiosperms typically possess scalariform perforations. There may be individual exceptions to most of these characters. Vessels are absent in some angiosperms (Winteraceae) while some gymnosperms have vessels (Gnetales).
The flowers are unisexual without perianth in several Amentiferae, which also exhibit anemo-phily. In spite of these and other exceptions, this combination of characters is unique to angiosperms and not found in any other group of seed plants.
What is the age of Angiosperms?
The time of origin of angiosperms is a matter of considerable debate. For many years, the earliest well-documented angiosperm fossil evidence was considered to be the form-genus Clavito-pollenites. Couper, (1958) described from Barremian and Aptian strata of Early Cretaceous of southern England (132 to 112 mya-million years), monosulcate pollen with distinctly sculptured exine, resembling the pollen of the extant genus Ascarina.
Brenner and Bickoff (1992) recorded similar but inaperturate pollen grains from the Valanginian (ca 135 mya) of the Helez formation of Israel, now considered to be the oldest record of angiosperm fossils (Taylor and Hickey, 1996).
The number and diversity of angiosperm fossils increased suddenly and by the end of the Early Cretaceous (ca 100 mya) period major groups of angiosperms, including herbaceous Magnoliidae, Magnoliales, Laurales, Winteroids, and Liliopsida were well represented.
In the Late Cretaceous, at least 50 percent of the species in the fossil flora were angiosperms. By the end of the Cretaceous, many extant angiosperm families had appeared. They subsequently increased exponentially and constituted the most dominant land flora, continuing up to the present.
The trail in the reverse direction is incomplete and confusing. Many claims of angiosperm records before the Cretaceous were made but largely rejected. Erdtman (1948) described Eucommiidites as a tricolpate dicotyledonous pollen grain from the Jurassic which had bilateral symmetry instead of the radial symmetry of angiosperms and granular exine with gymnospermous laminated endexine.
This pollen grain was also discovered in the micropyle of seeds of the female cone of uncertain but clearly gymnospermous affinities. Several other fossil pollens from the Jurassic age attributed to Nymphaeaceae ultimately turned out to be gymnosperms.
In the last few years fossils of Archaefructus from the Upper Cretaceous (nearly 124 mya) of China, with clearly defined spirally arranged conduplicate carpels enclosing ovules, a feature not reported in earlier angiosperms. The fruit is a follicle. This is considered to be the oldest record of the angiosperm flower.
Several vegetative structures from the Triassic were also attributed to angiosperms. Brown (1956) described Sanmiguilea leaves from the Late Triassic of Colorado and suggested an affinity with Palmae.
A better understanding of the plant was made by Cornet (1986, 1989), who regarded it as a presumed primitive angiosperm with features of monocots and dicots.
Although its angiosperm venation was refuted by Hickey and Doyle (1977), Cornet (1989) established its angiosperm venation and associated reproductive structures. Our knowledge of this controversial taxon, however, is far from clear.
Marcouia leaves (earlier described as Ctenis neuropteroides by Daugherty, 1941) are recorded from the Upper Triassic of Arizona and New Mexico. Its angiosperm affinities are not clear. Harris (1932) described Furcula from the Upper Triassic of Greenland as a bifurcate leaf with dichotomous venation.
Although it seems to approach dicots in venation and cuticular structure, it has several non-angiospermous characteristics including bifurcating midrib and blade, and higher vein orders with relatively acute angles of origin (Hickey and Doyle, 1977).
Cornet (1993) has described Pannaulika, a dicot-like leaf form from Late Triassic from the Virginia-North Carolina border.
It was considered to be a three-lobed palmately veined leaf. The associated reproductive structures were attributed to angiosperms but it is not certain that any of the reproductive structures were produced by the plant that bore Pannaulika.
What is the Place of their Origin?
It was previously believed that angiosperms originated in the Arctic region (Seward, 1931), and that they then migrated southward after their initial appearance. According to the upland theory proposed by Axelrod (1970), flowering plants first appeared in mild uplands located at low latitudes.
According to Smith (1970), the region of South-East Asia that is adjacent to Malaysia was the place where angiosperms emerged during the time that Gondwana and Laurasia were going through the first stages of their fragmentation.
Stebbins (1974) proposed that their ancestors originated in habitats that were open to the elements and located in regions that experienced seasonal dryness. Takhtajan (1966, 1980), who believed in the neotenous origin of angiosperms, suggested that angiosperms arose under environmental stress, probably as a result of adaptation to moderate seasonal draught on rocky mountain slopes in areas with monsoon climate.
According to Retallack and Dilcher’s (1981) theory, the first angiosperms were probably woody plants with little leaves that lived in the Rift valley system that connected Africa and South America. Some of these angiosperms adapted to coastal habitats and expanded as sea levels rose throughout the Early Cretaceous period.
While acknowledging the importance of environmental stress, numerous authors in recent years have proposed that early angiosperms lived near stream and lake edges (Hickey and Doyle, 1977; Upchurch and Wolfe, 1987; Hickey and Taylor, 1992). (lowland theory). They first appeared on river terraces, then later in the more secure back swamp and channel areas.
According to Taylor and Hickey (1996), the ancestors of angiosperms were perennial rhizomatous herbs that developed along rivers and streams at locations with moderate levels of alluviation and relatively high levels of disturbance. These areas would have had high nutrient levels and frequent plant cover loss as a result of recurring disturbances.
Are Angiosperms Monophyletic or Polyphyletic?
Engler (1892) considered angiosperms polyphyletic, monocotyledons, and dicotyledons as having evolved separately. The considerable diversity of angiosperms in the Early Cretaceous and the extant angiosperms led several authors, including Meeuse (1963) and Krassilov (1977), to develop models for the polyphyletic origin of angiosperms. This view is supported mainly by considerable diversity in the early angiosperm fossils.
The vast majority of contemporary researchers, such as Hutchinson (1959, 1973), Cronquist (1981, 1988), Thorne (1983, 1992, 2000, 2007), Dahlgren (1980, 1989), Takhtajan (1987, 1997), Judd et al. (2002, 2008), Bremer et al. (APG II, 2003), and Stevens (APweb, 2008), agree with the theory that angiosperms originated from a single common ancestor This perspective is backed by a unique combination of characters, some of which are as follows: closed carpels, sieve tubes, companion cells, four microsporangia, triploid endosperm, 8-nucleate embryo sac, and reduced gametophytes.
Sporne (1974), on the basis of statistical studies, also concluded that it is highly improbable that such a unique combination of characters could have arisen more than once, independently from gymnosperm ancestors.
It is interesting to note that Melville (1983) considered angiosperms to be monophyletic, but the explanation that he offers clubs him with the proponents of polyphyletic origin. He believes that angiosperms arose from several different genera of Glossopteridae.
According to him, the species is not always to be considered as the ancestor for determining a monophyletic nature. A species from another species is monophyletic, as is a genus from a genus, a family from a family. The principle, according to him, is that to be monophyletic, a taxon of any rank must be derived solely from another taxon of the same rank.
Glossopteridae and Angiospermidae belong to the same rank subclass. Both taxa consist of minor lineages that may be likened to a rope with many strands, a situation called pachyphyletic. This explanation, however, conforms to the concept of minimum monophyly and does not satisfy the rule of strict monophyly, which is now the accepted criterion for monophyly.
What are the possible ancestors of angiosperms?
The ancestry of angiosperms is perhaps one of the most controversial and vigorously debated topics. In the absence of direct fossil evidence, almost all groups of fossil and living gymnosperms have been considered possible ancestors.
Some authors even suggested the Isoetes origin of monocotyledons because the plant has a superficial resemblance with onion, although no trace of seed habit.
The various theories have revolved around two basic theories, viz., the Euanthial theory, and the Pseudanthial theory of angiosperm origin. Some other theories projecting herbaceous ancestry for the angiosperms have also recently received attention, making the question of ancestry of angiosperms rather more ambiguous:
Euanthial Theory
Also known as the Anthostrobilus theory, the Euanthial theory was first proposed by Arber and Parkins (1907). According to this theory, the angiosperm flower is interpreted as being derived from an unbranched bisexual strobilus bearing spirally arranged ovulate and pollen organs, similar to the hermaphrodite reproductive structures of some extinct Bennett Italian gymnosperms.
The carpel is thus regarded as a modified megasporophyll. The bisexual flower of Magnoliales has been considered to have evolved from such a structure.
Also agreeing with this general principle, various authors have tried to identify different gymnosperm groups as possible angiosperm ancestors:
Cycadeoidales (Bennettitales)
The group, now better known as Cycadeoidales, appeared in the Triassic and disappeared in the Cretaceous.
Their potential as angiosperm ancestors was largely built upon the studies of Wieland (1906, 1916). Lemesle (1946) considered the group to be ancestral to angiosperms, primarily because of the hermaphrodite nature of Cycadeoidea, which had an elongated receptacle with perianth-like bracts, a whorl of pollen-bearing microsporophylls surrounding the ovuliferous region having numerous.
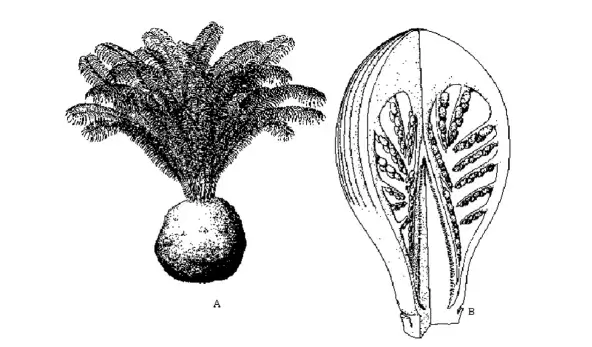
The plant was believed to look like cycads with a short compact trunk and a crown of pinnate compound leaves. It was earlier suggested that the microsporophylls opened at maturity but the subsequent studies of Crepet (1974) showed that microsporophylls were pinnate, and distal tips of pinnae were fused, the opening of the region was not structurally possible, and they later disintegrated internally.
The ovules were terminal in contrast to their position in carpels of angiosperms.
Caytoniaceae
Angiosperms are thought to have descended from Pteridosperms, also known as seed ferns. Pteridosperms are classified in the order Lyginopteridales, but Caytoniales are where they are most usually found. This theory has gained significant traction in recent years.
Thomas first identified the Caytoniaceae family from the Jurassic of Cayton Bay, which is located in Yorkshire. Subsequent descriptions came from Greenland, England, and Canada. The group first appeared in the Late Triassic and went extinct in the Cretaceous Period towards the end of that period.
The leaves (Sagenopteris) grew on twigs rather than the trunk. These were net-veined and had two pairs of leaflets (rarely three to six leaflets). Caytonanthus male structures had rachis with branching pinnae, each with a synangium of four microsporangia. Caytonia’s seed-bearing structure had a rachis with two rows of stalked cupules (Figure 2).
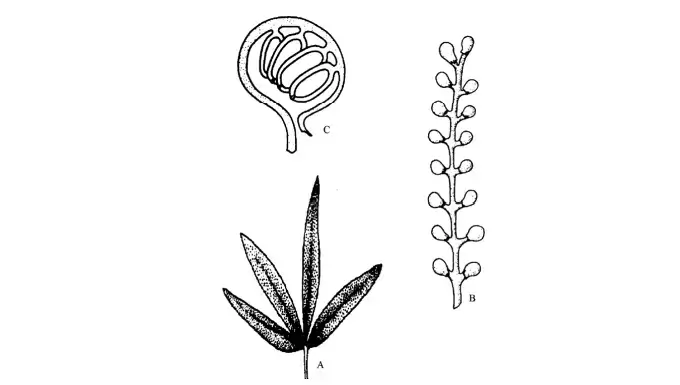
Each cupule contained several ovules that were borne in such a way that the cupule was recurved, with a lip-like projection (often referred to as the stigmatic surface) near the point of attachment (Fig 3).
The discovery of pollen grains within the ovules was thought to indicate that they were gymnosperm ancestors rather than angiosperm ancestors.
Krassilov (1977) and Doyle (1978) considered the cupule to be homologous to the carpel, whereas Gaussen (1946) and Stebbins (1974) considered it to be the ovule’s outer integument. Doyle and Donoghue’s (1987) cladistic studies support the caytoniales-angiosperm lineage. Thorne (1996) agreed that angiosperms most likely evolved from seed ferns during the Late Jurassic.
Cycadales:
Sporne (1971) suggested possible links between Cycadales and angiosperms in the palm-like habit of Cycadales. The ovules are borne on leaf-like megasporophylls.
Trends in the reduction of sporophyll blade as seen in various species of Cycas. Although it may be difficult to assume Cycadales as ancestral to angiosperms.
Pseudanthium Theory
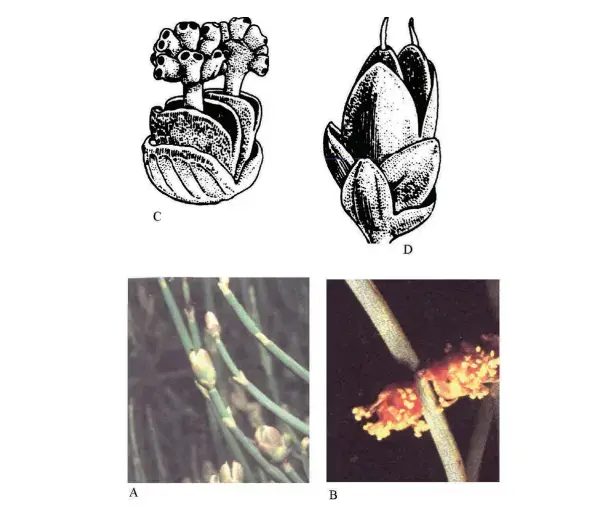
Commonly associated with the Englerian School, the theory was first proposed by Wettstein (1907), who postulated that angiosperms were derived from the Gnetopsida, represented by Ephedra, Gnetum, and Welwitschia (formerly all placed in the same order Gnetales).
The group shows more angiosperm characteristics than any other group of living or fossil gymnosperms. These include the presence of vessels, reticulate dicot-like leaves (Gnetum), male flowers with perianth and bracts, strong gametophyte reduction, and fusion of the second male gametophyte with the ventral canal nucleus.
Ephedra resembles Casuarina inhabit. Wettstein homologized the compound strobili of Gnetales with the inflorescences of wind-pollinated Amentiferae. Showy insect-pollinated bisexual flowers of Magnolia as pseudanthia derived by aggregation of unisexual units, the carpel thus representing a modified branch (Stachyosporous origin of carpel).
Gnetopsida is a very young group. But this theory has been strongly supported by Young (1981), who challenged the view that first angiosperms were vessel-less and assumed that vessels were lost in several early lines.
Muhammad and Sattler (1982) found scalariform perforations in vessel elements of Gnetum, suggesting that angiosperms may be derived from Gnetales after all. Carlquist (1996), however, concludes that this claim from Gnetum does not hold when large samples are examined.
According to this theory, the amentiferous-hamamelid orders Casuarinales, Fagales, Myricales, and Juglandales made constituted the most fundamental group of angiosperms. It is important to note that Wettstein (1907) includes Chloranthaceae and Piperaceae in this fundamental group.
These families have attracted a significant portion of interest in recent years, thus Wettstein’s mention of them is significant. The discovery of a fossil Welwitschia-like plant from the Late Triassic of Texas, identified by Cornet (1996) as Archaestrobilus cupulanthus, has further strengthened the significance of Gnetopsids in angiosperm phylogeny (Figure 4).
The plant’s male and female spikes, which each contained hundreds of spirally arranged macrocupules, were built similarly. While female spikes appeared singly, male spikes were borne in groups of three. Each female macrocupule was axially symmetric.

Ephedra is generally considered to be the most primitive of the three living genera of gnetopsids. Cornet believes that Archaestrobilus possessed characters that may be plesiomorphic even for Ephedra.
Anthocorm Theory
This theory is a modified version of the pseudanthial theory and was proposed by Neumayer (1924). According to this theory, the angiosperm flower has several separate origins (i.e. angiosperms are polyphyletic). In most Magnoliidae and their dicotyledonous derivatives, they are modified paraxial systems (holanthocorms). They originated from the gnetopsids through the Piperales.
whereas the modification of an originally uniaxial system (gonoclad or anthoid) gave rise to flowers of Chloranthaceae. Meeuse (1963) postulated a separate origin of monocotyledons from the fossil order Pentoxylales through the monocot order Pandanales.
Gonophyll Theory
The Gonophyll theory was developed by Melville, largely on the basis of a study of the venation pattern. He derived angiosperms from Glossopteridae, which formed important elements in the flora of Gondwanaland. He further derived angiosperm flower from gonophyll (a fertile branching axis adnate to a leaf).
Glossopterids In Scutum and Ottokaria, the fertile branch was composed of a bivalved scale known as the scutella, which had terminal ovules on dichotomous groups of branches. A hint of this closure can be found in the Permian fossil Breytenia. The angiosperm condition is developed by folding the scutella along the cluster of its ovules to establish the condition of an angiosperm.
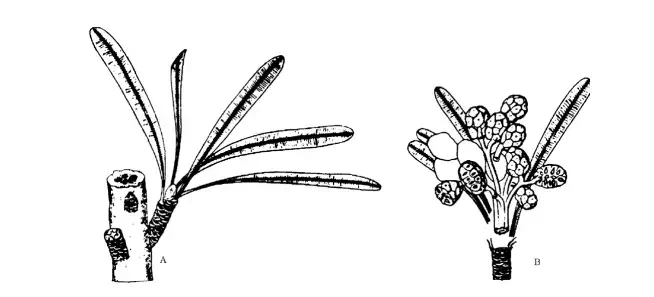
The fertile branch of a Lidgettonia comprises four to eight disc-like structures, each of which bears multiple seeds. Approximately six seed-bearing cupules are linked to long stalks borne from the midrib of fertile scale in the species.

Herbaceous Origin Hypothesis
The herbaceous origin hypothesis is similar to the Pseudanthial theory; however, the herbaceous origin hypothesis suggests that the ancestor plant was a perennial rhizomatous herb rather than a tree.
The term “paleoherb” was first used by Donoghue and Doyle (1989) to refer to a group of derivative (not ancestral) forms of the Magnoliidae family that had anomocytic stomata, two whorls of the perianth, and trimerous flowers. This group included the Lactoridaceae, Aristolochiaceae, Cabombaceae, Piperales, Nymphaeaceae
This theory proposes that the ancestors of angiosperms were small, herbaceous plants that had a rhizomatous to scrambling perennial growth habit. They had simple leaves that were veined in a reticulated pattern and had a primary venation pattern that could have been palmate or pinnate, but the secondary veins branched in a dichotomous manner.
Sieve-tube elements and elongated tracheary elements with circular-bordered and scalariform pitting and oblique end walls were present in the vegetative anatomy. Inflorescences that ranged from cymose to racemose contained the flowers.
The monosulcate pollen grains were perforated, giving them a reticulate sculptured appearance. The carpels were free and ascidiate, which means that the ovules were attached proximally to the closure. Each carpel contained one or two orthotropous, bitegmic, crassinucellate ovules as well as a dicotyledonous embryo.
The aforementioned authors note that fossil angiosperm wood is extremely rare, whereas early fossils typically contain a large number of leaf impressions.
The consensus is emerging from recent phylogenetic studies that gnetopsida represents the closest living relatives of angiosperms, whereas the closest fossil group is the Bennettitales. Angiosperm lineage, together with these two groups, constitutes Anthophytes. The group is believed to have split in the late Triassic.
The angiosperm lineage continued as Angiophytes up to the Late Jurassic when it further split into stem Angiophytes and crown Angiophytes.
Transitional-Combinational Theory
In 2004, Stuessy published a transitional-combinational theory for the origin of angiosperms. This publication brought a new interest in angiosperms and explained several recent viewpoints and discoveries that were in conflict with one another.
According to this theory, angiosperms evolved slowly from seed ferns during the Jurassic period. The development of the carpel came first, then came the process of double fertilization, and finally came the formation of the flower. It’s possible that more than 100 million years passed between each of these three significant phases of evolution.
The theory is put forward in light of the fact that it is difficult to locate the ancestors of angiosperms, but also in light of the fact that they suddenly appeared and experienced rapid evolutionary success.
According to the fossil record, the first known occurrence of angiosperms did not take place until the Early Cretaceous epoch, which was when the final combination of the three most important angiosperm characteristics took place.
This combination opened the door to an explosion of evolutionary diversity, particularly as a result of selection by insect pollinators, as well as the accompanying modifications in compatibility and breeding systems.
This theory attempts to explain the discrepancy between fossil and molecular phylogenetic data, the latter of which suggests that the origin of angiosperms occurred prior to the Cretaceous period when DNA (and protein) sequences showed the first changes accompanying the evolution of carpels, which occurred much earlier than the final combination of all three angiosperm characteristics.
In addition, the theory suggests that, with the exception of extinct seed ferns, from which the carpel evolved, other gymnosperms had no direct phylogenetic ties to modern angiosperms.
Origin of Monocotyledons
At first, it was thought (Engler, 1892; Meeuse, 1963) that monocotyledons came before dicotyledons and are polyphyletic. Most people now think that monocotyledons evolved from dicots in a single step. Bailey (1944) and Cheadle (1953) said that the origin and specialization of vessels in monocots and dicots were different, so dicots without vessels gave rise to monocots.
Cronquist did not agree that vessels in two groups came from different places. He thought that monocots came from aquatic ancestors that looked like modern Nymphaeales. Kosakai et alstudies .’s of ships, on the other hand, showed that this was not true (1970).
They found it hard to believe that supposedly primitive Alismataceae evolved advanced vessels in an aquatic environment and then gave rise to terrestrial monocots with more primitive vessel elements in the metaxylem of their roots. So, they thought it was more likely that Alismataceae came from plants that lived on land.
Hutchinson (1973) says that monocots developed from Ranales along two lines, with Ranunculoideae leading to Alismatales and Helleboroideae leading to Butomales. Takhtajan (1980, 1987) suggested that both the Nymphaeales and the Alismatales came from a group of herbaceous land plants called Magnoliidae.
Dahlgren et al. (1985) thought that monocots first appeared in the Early Cretaceous, about 110 million years ago. At that time, the ancestors of Magnoliiflorae must have already had some of the characteristics of this group, but they were not as different from each other as they are now. Other dicotyledonous groups had already split off from the ancestral stock.
Thorne (1996) says that monocotyledons seem to have come from the most primitive dicotyledons very early on. Chase et al. (1993) and Qiu et al. (1993) looked at the rbcL sequences of plants and found that the monocots were monophyletic and came from the monosulcate Magnoliidae. The least specialized monocotyledons are thought to be Acorus, Melanthiaceae, and Butomus.
Questions about the Origin of Angiosperms:
Where did angiosperms originate from?
Angiosperms are plants that produce flowers that may include either male or female reproductive organs. These plants are often known as “seed in a vessel” plants. Evidence from fossils suggests that flowering plants initially appeared in the Lower Cretaceous, which occurred approximately 125 million years ago, and that by the Middle Cretaceous, which occurred approximately 100 million years ago, flowering plants were evolving rapidly.
What is the ancestor of angiosperms?
Gymnosperms and angiosperms evolved from a common ancestor that lived around 300 million years ago. This ancestor was some form of seed fern, a plant that bore seeds and pollen on its leaves.
Which group of plants evolved first?
More than 700 million years ago, the first plants were algae living in the oceans. These aquatic algae with no stems or roots gave rise to modern plants. Plant evolution on land has been ongoing throughout geologic time.
What is Gonophyll
The basic component of the angiosperm ovary, according to Melville (1963), was a leaf with an epiphyllous fertile branch called the gonophyll. The evidence pointed to the ovary being made up of alternating whorls of foliar and fertile organs.
What are the possible ancestors of angiosperms?
The ancestry of angiosperms is perhaps one of the most controversial and vigorously debated topics. In the absence of direct fossil evidence, almost all groups of fossil and living gymnosperms have been considered possible ancestors. Some authors even suggested the Isoetes origin of monocotyledons because the plant has a superficial resemblance with onion, although no trace of seed habit.
What is the age of Angiosperms?
The time of origin of angiosperms is a matter of considerable debate. For many years, the earliest well-documented angiosperm fossil was considered to be the form-genus Clavito-pollenites. Couper, (1958) described from Barremian and Aptian strata of the Early Cretaceous of southern England (132 to 112 mya-million years), monosulcate pollen with distinctly sculptured exine, resembling the pollen of the extant genus Ascarina.
Sources and References:
- Google. (n.d.). Plant Systematics. Google Books. Retrieved July 12, 2022, from This Link
- Fig. Flowering Orchid Available Here
- https://en.wikipedia.org/wiki/Evolutionary_history_of_plants