
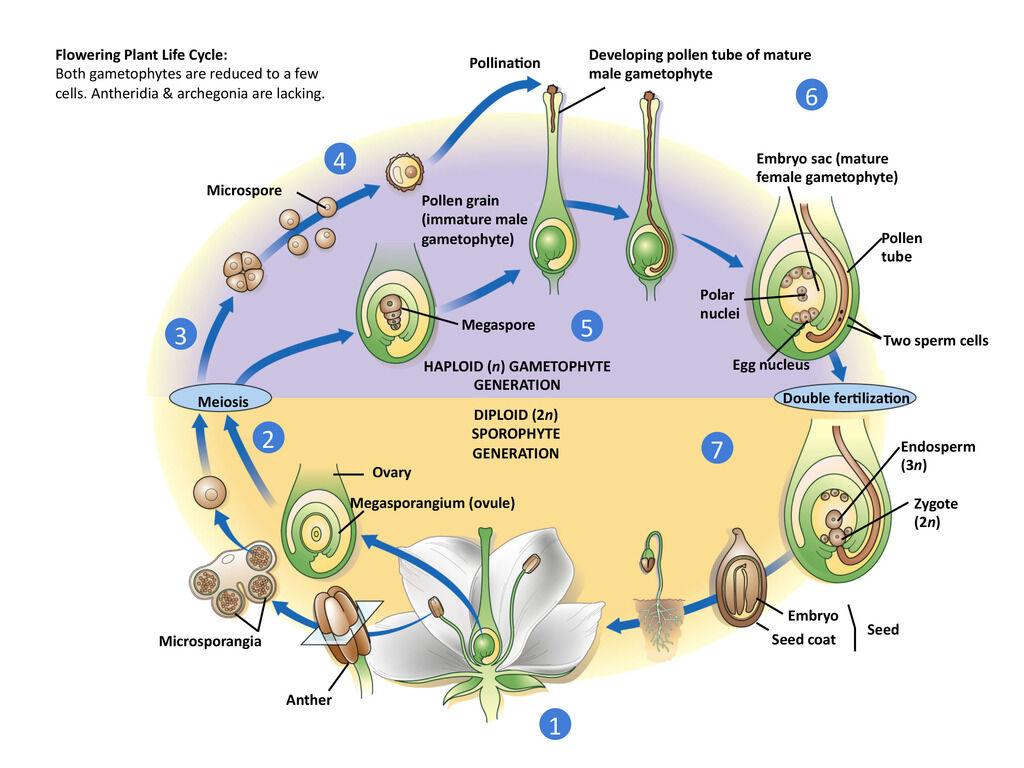
During the development of a flower bud, a single megaspore mother cell in the ovule undergoes meiosis, producing four mega-spores (figure ). In most flowering plants, three of the megaspores soon disappear; the nucleus of the remaining mega-spore divides mitotically, and the cell slowly expands until it becomes many times its original size.
The female gametophyte
While the expansion of the megaspore is occurring, each of the daughter nuclei divides twice, resulting in eight haploid nuclei arranged in two groups of four. At the same time, two layers of the ovule, the integuments, differentiate and become the seed coat of a seed. The integuments, as they develop, form the micropyle, a small gap or pore at one end.
Pollen production
While the female gametophyte is developing, a similar but less complex process takes place in the ant-hers (see figure ). Most anthers have patches of tissue (usually four) that eventually become chambers lined with nutritive cells.
The tissue in each patch is composed of many diploid microspore mother cells that undergo meiosis more or less simultaneously, each producing four microspores.
The four micros-pores at first remain together as a quartet or tetrad, and the nucleus of each microspore divides once; in most species, the microspores of each quartet then separate. At the same time, a two-layered wall develops around each microspore.
As the microspore-containing anther continues to mature, the wall between adjacent pairs of chambers breaks down, leaving two larger sacs.
At this point, the binucleate microspores have become pollen grains. The outer pollen grain wall layer often becomes beautifully sculptured, and it contains chemicals that may react with others in a stigma to signal whether the development of the male gametophyte should proceed to completion. The pollen grain has areas called apertures, through which a pollen tube may later emerge.
Pollination and the male gametophyte
Pollination is simply the mechanical transfer of pollen from its source (an anther) to a receptive area (the stigma of a flowering plant). Most pollination takes place between flowers of different plants and is brought about by insects, wind, water, gravity, bats, and other animals.
In as many as one-quarter of all angiosperms, however, a pollen grain may be deposited directly on the stigma of its own flower, and self-pollination occurs.
Pollination may or may not be followed by fertilization, depending on the genetic compatibility of the pollen grain and the flower on whose stigma it has landed. If the stigma is receptive, the pollen grain’s dense cytoplasm absorbs substances from the stigma and bulges through an aperture. The bulge develops into a pollen tube that responds to chemical and mechanical stimuli that guide it to the embryo sac.
It follows a diffusion gradient of the chemicals and grows down through the style and into the micropyle. The pollen tube usually takes several hours to two days to reach the micropyle, but in a few instances, the journey may take up to a year.
Pollen tube growth is more rapid in angiosperms than in gymnosperms. Rapid pollen tube growth rate is an innovation that is believed to have preceded fruit development and been essential in the origins of angiosperms (see figure ).
One of the pollen grain’s two cells, the generative cell, lags behind. Its nucleus divides in the pollen grain or in the pollen tube, producing two sperm cells. Unlike sperm in mosses, ferns, and some gymnosperms, the sperm of flowering plants have no flagella. At this point, the pollen grain with its tube and sperm has become a mature male gametophyte.
Double fertilization and seed production
As the pollen tube enters the embryo sac, it destroys a synergid in the process and then discharges its contents. Both sperm are functional, and an event called double fertilization follows. One sperm unites with the egg and forms a zygote, which develops into an embryo sporophyte plant. The other sperm and the two polar nuclei unite, forming a triploid primary endosperm nucleus.
The primary endosperm nucleus begins dividing rapidly and repeatedly, becoming triploid endosperm tissue that may soon consist of thousands of cells. Endosperm tissue can become an extensive part of the seed in grasses such as corn, and it provides nutrients for the embryo in most flowering plants.
Until recently, the nutritional, triploid endosperm was believed to be the ancestral state in angiosperms. A recent analysis of extant, basal angiosperms revealed that diploid endosperms were also common.
The female gametophyte in these species has four, not eight nuclei. At the moment, it is unclear whether diploid or triploid endosperms are the most primitive.
Germination and growth of the sporophyte
As mentioned earlier, a seed may remain dormant for many years, depending on the species. When environmental conditions become favorable, the seed undergoes germination, and the young sporophyte plant emerges.
Again depending on the species, the sporophyte may grow and develop for many years before becoming capable of reproduction, or it may quickly grow and produce flowers in a single growing season.
References:
www.ravenbiology.com
https://sciencedocbox.com
https://onlinelibrary.wiley.com